
Поперечнополосатая мышечная ткань Различают поперечнополосатую скелетную и сердечную мышечные ткани. Поперечнополосатая скелетная мышечная ткань Состоит из продольно ориентированных
Сердечная мышечная ткань. Строение сердечной мышечной ткани.
Гладкая мышечная ткань мочевого пузыря 1 — гладкие миоциты (клетки, образующие гладкую мышечную ткань): имеют веретиновидную форму и часто объединяются в пучки. Толстые и тонкие миофиламенты
Поперечнополосатая мышечная ткань языка кролика гистология
Сердечная мышечная ткань. Строение сердечной мышечной ткани.
Гистогенез сердечной мышечной ткани. Источники развития сердечной мышечной ткани находятся в прекардиальной мезодерме. В гистогенезе возникают парные складчатые утолщения висцерального листка спланхнотома — миоэпикардиальные пластинки, содержащие стволовые клетки сердечной мышечной ткани. Последние путем дивергентной дифференцировки дают начало следующим клеточным дифферонам: рабочим, ритмзадающим (пейсмекерным), проводящим и секреторным кардиомиоцитам.
Исходные клетки сердечной мышечной ткани — кардиомиобласты характеризуются рядом признаков: клетки уплощены, содержат крупное ядро, светлую цитоплазму, бедную рибосомами и митохондриями. В дальнейшем происходит развитие комплекса Гольджи, гранулярной эндоплазматической сети. В кардиомиобластах обнаруживаются фибриллярные структуры, но миофибрилл нет. Клетки обладают высоким пролиферативным потенциалом. После ряда митотических циклов кардиомиобласты дифференцируются в кардиомиоциты, в которых начинается саркомерогенез. В цитоплазме кардиомиоцитов увеличивается число полисом, канальцев гранулярной эндоплазматической сети, накапливаются гранулы гликогена, возрастает объем актомиозинового комплекса. Кардиомиоциты сокращаются, но не теряют способность к дальнейшей пролиферации и дифференцировке. Развитие сократительного аппарата в позднем эмбриональном и постнатальном периодах происходит путем надставки новых саркомеров и наслоения вновь синтезированных миофиламентов. Дифференцировка кардиомиоцитов сопровождается увеличением числа митохондрий, распределением их у полюсов ядер и между миофибриллами и протекает параллельно со специализацией контактирующих поверхностей клеток. Кардиомиоциты путем контактов «конец в конец», «конец в бок» формируют клеточные комплексы — сердечные мышечные волокна, и в целом ткань представляет собой сетевидную структуру.
Строение сердечной мышечной ткани.
Структурно-функциональные единицы волокон — кардиомиоциты — это клетки, имеющие вытянутую прямоугольную форму. Длина рабочих кардиомиоцитов составляет 50-120 мкм, а ширина — 15-20 мкм. Одно-два ядра располагаются в центре клетки. Периферическую часть цитоплазмы кардиомиоцитов занимают поперечноисчерченные миофибриллы, аналогичные таковым в симпластах скелетномышечного волокна. Однако каналы саркоплазматической сети и Т-системы менее отчетливо выражены. Кардиомиоциты отличаются большим количеством митохондрий, расположенных тесными рядами между миофибриллами. Снаружи миоциты покрыты сарколеммой, в составе которой выделяются плазмолемма и базальная мембрана. Характерной особенностью ткани является наличие вставочных дисков на границе между контактирующими кардиомиоцитами. Вставочные диски пересекают волокно в виде волнистой или ступенчатой линии и включают межклеточные контакты от простых, по типу десмо-сом и до щелевых (нексусов).
Часть кардиомиоцитов на ранних этапах кардиомиогенеза являются сократительно-секреторными. В дальнейшем в результате дивергентной дифференцировки возникают «темные» (сократительные) и «светлые» (проводящие) миоциты, в которых исчезают секреторные гранулы, тогда как в предсердных миоцитах они сохраняются. Так формируется дифферон эндокринных кардиомиоцитов. Эти клетки содержат центрально расположенное ядро с диспергированным хроматином,
1-2 ядрышками. В цитоплазме хорошо развиты гранулярная эндоплазматическая сеть, диктиосомы комплекса Гольджи, в тесной связи с элементами которого находятся многочисленные секреторные гранулы диаметром около 2 мкм, содержащие электронноплотный материал. В дальнейшем секреторные гранулы обнаруживаются под сарколеммой и выделяются в межклеточное пространство путем экзоцитоза. Выделенный пептидный гормон кардиодилатин циркулирует в крови в виде кардионатрина, который вызывает сокращение гладких миоцитов артериол, увеличение почечного кровотока, ускоряет клубочковую фильтрацию и выделение натрия из организма.
Кардиомиоциты проводящей системы гетероморфны. В них слабо развит мио-фибриллярный аппарат, расположение миофиламентов в составе миофибрилл рыхлое, Z-линии имеют неправильную конфигурацию, эндоплазматическая сеть слабо развита, находится на периферии миоцитов, число митохондрий незначительное. По мере расположения этих кардиомиоцитов в проксимо-дистальном направлении соответственно движению импульсов от синусно-предсердного узла, через предсердно-желудочковый узел, пучок Гиса, его ножки и клетки Пуркиня к рабочим миоцитам проводящие кардиомиоциты по своей ультраструктуре приближаются к рабочим кардиомиоцитам.
Регенерация сердечной мышечной ткани.
В гистогенезе сердечной мышечной ткани специализированный камбий не возникает. Поэтому регенерация ткани протекает на основе внутриклеточных гиперпластических процессов. Вместе с тем для кардиомиоцитов млекопитающих, приматов и человека характерен процесс полиплоидизации. Например, у обезьян ядра до 50% терминально дифференцированных кардиомиоцитов становятся тетра- и октоплоидными. Полиплоидные кардиомиоциты возникают за счет ацитокинетического митоза, что приводит к многоядерности.
В условиях патологии сердечно-сосудистой системы человека (ревматизм, врожденные пороки сердца, инфаркт миокарда и другие) важная роль в компенсации повреждений кардиомиоцитов принадлежит внутриклеточной регенерации, полиплоидизации как ядер, так и кардиомиоцитов.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Мышечные ткани
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
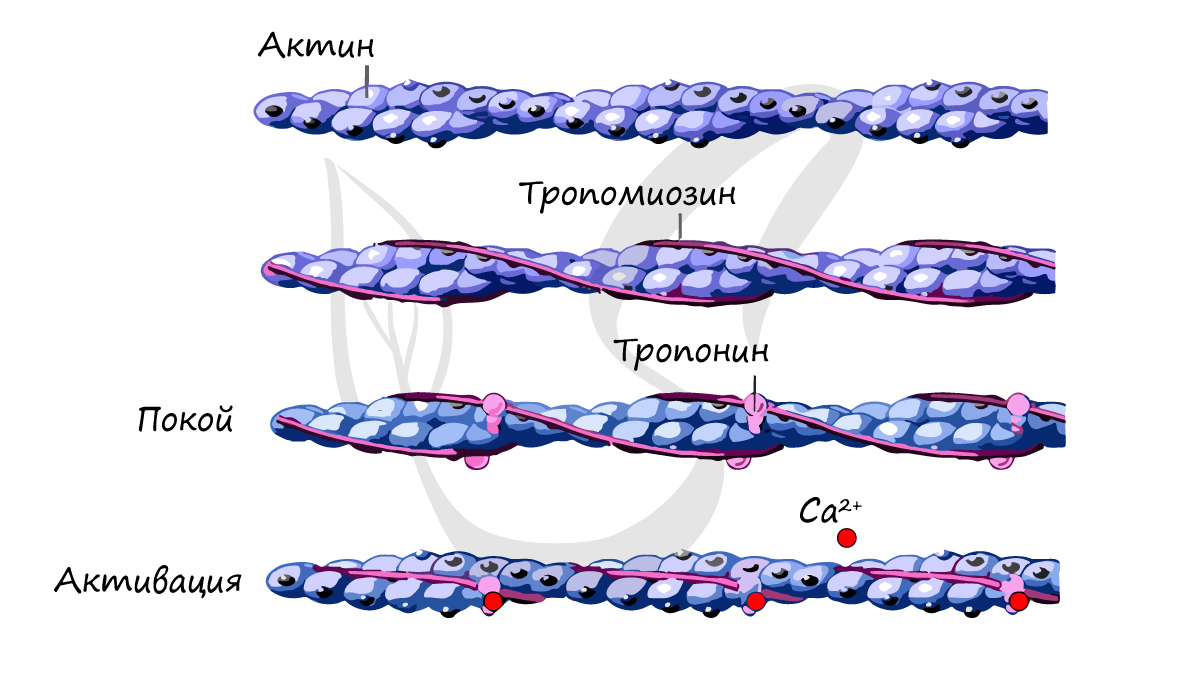
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
МЫШЕЧНАЯ ТКАНЬ
МЫШЕЧНАЯ ТКАНЬ (textus muscularis, LNH) — группа тканей организма животных и человека, обладающих свойством сократимости. Выделяют гладкую и поперечнополосатую Мышечная ткань; последнюю, в свою очередь, подразделяют на скелетную и сердечную. Свойством сократимости обладают также некоторые специализированные разновидности других тканей — миоэпителиальная ткань (эктодермального происхождения), входящая в состав концевых секреторных отделов (ацинусов) слюнных и потовых желез, и Мышечную ткань нейроглиального происхождения, являющаяся составной частью радужки (сфинктер и дилататор зрачка).
В разработку вопроса о природе и происхождении Мышечной ткани большой вклад внесли А. А. Заварзин, Н. Г. Хлопин и другие отечественные ученые. Гладкая М. т. человека и позвоночных животных развивается в составе производных спланхноплевры — висцерального листка спланхнотомов (см. Мезодерма) и по своей природе относится к тканям внутренней среды организма. Скелетная М. т. возникает из специфического эмбрионального зачатка — миотома, входящего в состав элементов сегментированной мезодермы — сомитов (см.), а также из мезодермального зачатка — мезенхимы (см.). Сердечная М. т. развивается из целомического эпителия. Для всех М. т. характерно сходное обособление в составе эмбрионального зачатка в виде клеток веретенообразной формы — мышцеобразовательных клеток, или миобластов.
Содержание
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ
Гладкая М. т. (textus muscularis nonstriatus) человека и позвоночных животных формирует мышечные оболочки внутренних органов и стенки кровеносных сосудов.
Гистогенез
Для эмбрионального гистогенеза гладкой М. т. характерно обособление среди мезенхимы, окружающей развивающийся орган или сосуды (кишечную трубку, трахею, бронхи, мочеточники, аорту, артерии, вены и т. д.), веретенообразных клеток, располагающихся обычно в два слоя (внутренняя и наружная мышечные оболочки). В процессе дифференцировки гладкой М. т. гладкие мышечные клетки — гладкие миоциты (myocytus glaber), или лейомиоциты, сближаются и образуют плотную структуру, приобретая специфическую дефинитивную форму.
Строение

Рис. 1. Микропрепараты мышечной оболочки кишечника саламандры (продольный разрез). Гладкая мышечная ткань в расслабленном (а) и сокращенном (б) состоянии: 1 — ядра; 2 — миофибриллы.
В мышечной оболочке большинства внутренних органов гладкие мышечные клетки имеют удлиненную (от 15 до 500 мкм) веретенообразную форму. В соответствии с формой клетки ядра вытянуты в длину, причем у млекопитающих при сокращении клетки ядро может приобретать штопорообразную форму. У нек-рых видов амфибий сокращение гладкой мышечной клетки сопровождается сжатием ядра вдоль оси (рис. 1). Митохондрии (см.) в гладкой мышечной клетке концентрируются вокруг ядра, а также вдоль оси клетки. Гладкая и шероховатая эндоплазматическая сеть (см. Эндоплазматический ретикулум) развита слабо.

Рис. 2. Объемная схема ультраструктуры участка гладкой мышечной ткани позвоночных: а — мышечные клетки; б — небольшой участок клеток, изображенных на схеме а (1 — ядра; 2 — зона тесного контакта клеток; 3 — митохондрии; 4 — плазмолемма; 5 — впячивание плазмолеммы; 6 — эндоплазматическая сеть; 7 — протофибриллы).
По данным электронной микроскопии, ультраструктура гладких мышечных клеток характеризуется наличием многочисленных плазмо-леммальных виячиваний типа пиноцитозныхпузырьков (рис. 2). Предполагают, что с этими впячиваниями связана передача внутрь клетки раздражения, вызывающего ее возбуждение и сокращение.
Специфическим структурным признаком гладкой мышечной клетки, обнаруживаемым методом обычной световой микроскопии, является наличие в цитоплазме тонких волокон — миофибрилл, хорошо различимых на окрашенных железным гематоксилином микропрепаратах. На электронограммах выявляется, что миофибриллы состоят из более тонких волоконец — протофибрилл, или миофиламентов. Между клетками располагается строма гладкой М. т.— коллагеновые (ретикулярные) и эластические волокна, образующие плотные сети вокруг каждой клетки. Доказано, что гладкие мышечные клетки синтезируют волокна стромы при помощи шероховатой эндоплазматической сети.
Процесс сокращения в гладкой М. т. на обычных гистол, препаратах, окрашенных железным гематоксилином, обнаруживается по характерным уплотнениям (полосам сокращения), проходящим через многие ряды клеток. На электронограммах сокращение гладких мышечных клеток выявляется сгущением протофибрилл.
ПОПЕРЕЧНОПОЛОСАТАЯ МЫШЕЧНАЯ ТКАНЬ
Поперечнополосатая мышечная ткань (textus muscularis transverso-striatus) представлена скелетной и сердечной М. т.

Рис. 3. Схема процесса дифференцировки поперечнополосатых мышечных волокон в эмбриогенезе: а — стадия миобластов (часть миобластов находится в стадии митоза); б — слияние миобластов в мышечные трубочки (указаны стрелкой); в — сформированная мышечная трубочка; г — мышечное волокно: 1—центрально-осевой пучок миофибрилл, 2 — ядра мышечного волокна.
Скелетная мышечная ткань
Скелетная мышечная ткань [textus muscularis striatus (sceleti)]. Гистогенез. Основной источник развития скелетной М. т. в эмбриогенезе — мезодерма сомитов, из к-рой возникают «текучие» (перемещающиеся от места закладки к скелетным частям) закладки, состоящие из мезенхимных клеток, формирующих первичные модели мышц на месте их дифференцировки. Дифференцирующаяся мезенхимная клетка принимает веретенообразную форму миобласта, характеризующую первый (клеточный) этап дифференцировки. Число миобластов увеличивается за счет митотических делений, а также, как предполагают, путем превращения клеток-сателлитов (немногочисленные одноядерные клетки с уплотненным расположением хроматина в ядре и слабым развитием цитоплазматических органелл). При переходе к следующему, второму этапу дифференцировки митотические деления прекращаются и миобласты сливаются концами, образуя мышечные трубочки (myotubuli), в к-рых ядра занимают центрально-осевое положение (рис. 3). В периферической зоне мышечных трубочек формируются фибриллярные и мембранные структуры. Дифференцировка миобластов и мышечных трубочек может происходить и в культуре ткани вне организма. Однако третий этап дифференцировки — превращение мышечных трубочек в мышечные волокна с периферическим расположением ядер и центрально-осевым расположением миофибрилл может происходить только в организме в результате взаимодействия с двигательными нервными волокнами (см. Мышцы).
Строение

Рис. 4. Классическая схема строения мышечного волокна по Гейденгайну: 1 — диск А; 2 — диск I; 3 — перегородка T (телофрагма); 4 — перегородка М (мезофрагма); 5 — саркосомы; 6 — поперечная сеть; 7 — миофибриллы.

Рис. 5. Электронограмма саркомера: 1 — саркомер; 2 — диск А; 3 — полудиски; 4 — телофрагма; 5 — мезофрагма; 6 — толстые (миозиновые) миофиламенты; 7 — тонкие (актиновые) миофиламенты.
Скелетная М. т. построена из удлиненных многоядерных образований — поперечнополосатых мышечных волокон (туо-fibra transversostriata), каждое из к-рых имеет форму цилиндра с округленными или заостренными концами, средние размеры волокон (у млекопитающих и человека) — до 80 мкм в диаметре и до 12 см в длину. Как правило, у позвоночных каждое волокно составляет одиночную нить. Характерная особенность поперечнополосатых мышечных волокон, определяемая с помощью светового микроскопа,— поперечная исчерченность, обусловленная чередованием полосок, или дисков, двулучепреломляющего (анизотропного) вещества — диски A [stria А (discus А)] и лишенного двулучепре-ломления (изотропного) вещества — диски I [stria I (discus I)] (рис. 4). Диски А и I входят в состав расположенных вдоль оси поперечнополосатого мышечного волокна тонких волоконец — миофибрилл (myofibril-lae transversostriatae), образуя так наз. саркомеры (рис. 5). Границы саркомеров имеют вид тонких перегородок, пересекающих диски I в поперечном направлении, — телофрагм (telophragma), или перегородок Т; по старой терминологии линия Z. Менее четко выявляется проходящая поперек диска А перегородка — мезофрагма (mesophragma); по старой терминологии — линия М. У амфибий, рептилий, птиц, млекопитающих и человека миофибриллы на поперечных срезах имеют очертания круга или многоугольника диам. 1—2 мкм. У многих видов рыб, а также у членистоногих, для к-рых характерна поперечнополосатая М. т., миофибриллы отличаются уплощенной формой и на поперечных срезах имеют вид удлиненных овалов. Поперечнополосатое мышечное волокно можно рассматривать как гигантскую многоядерную клетку, цитоплазма которой (называемая обычно саркоплазмой) оттеснена к периферии занимающими центрально-осевое положение миофибрилла-ми. У всех позвоночных уплощенные ядра эллипсоидальной формы располагаются по периферии волокна (для костистых рыб и членистоногих характерно центральноосевое расположение ядер).
Под базальной мембраной мышечных волокон на электронограммах обнаруживают клетки-сателлиты.

Рис. 6. Микропрепарат мышечной ткани языка саламандры (поперечнополосатые мышечные волокна): а — волокно в расслабленном состоянии; б — волокно в начале сокращения (1 — ядро; 2 — диск А; 3 — диск I; 4 — перегородка T (телофрагма); 5 — диск А в фазе расщепления на два полудиска; 6 — полоска L).
Процесс сокращения поперечнополосатых мышечных волокон при исследовании под световым микроскопом характеризуется укорочением и утолщением саркомеров, что сопровождается перераспределением анизотропного и изотропного веществ (рис. 6). Характерная морфол, черта динамики сокращения саркомера заключается в расщеплении диска А на два полудиска, между к-рыми появляется светлая полоска Н, или светлая зона (stria L, zona lucida). По мере сокращения саркомера анизотропное вещество расходится к сближающимся телофрагмам, образуя в результате этого движения полосы сокращения, замещающие собой диски I.

Рис. 7. Объемная схема ультраструктуры телофрагмы: тонкие миофиламенты (1) сходятся концами (2), образуя заостренные на вершинах арки.
При электронно-микроскопическом исследовании поперечнополосатых мышечных волокон были выявлены тонкие детали строения миофибрилл и открыты слабо различимые при светооптическом исследовании мембранные структуры. Выяснилось, что миофибриллы поперечнополосатой М. т. состоят из ультраструктурных волоконец — протофибрилл, или миофиламентов, двух типов: толстых (ок. 11 нм в поперечнике) и тонких (ок. 5 нм). В составе саркомера толстые протофибриллы располагаются в зоне диска А, тонкие — в зоне диска I, образуя два полудиска, разделенные телофрагмой, к-рая построена концами тонких протофибрилл, сходящимися в виде заостренных арок (рис. 7). С помощью специфических биохим, методов исследования установлено, что протофибриллы построены из особых мышечных белков — толстые из миозина, тонкие в основном из актина. Строение молекул мышечных белков — см. ниже, раздел Биохимия мышечной ткани.

Рис. 8. Мембранный аппарат поперечнополосатых мышечных волокон: 1 — поперечные срезы миофибрилл; 2 — сарколемма; 3 — трубочка Т-системы; 4 — цистерны саркоплазматического ретикулума; 5 — канальцы саркоплазматического ретикулума (L-канальцы); 6 — переплет L-канальцев; 7 — триады; 8 — митохондрии; 9 — тонкие миофиламенты; 1 о — толстые миофиламенты.
Мембранный аппарат поперечнополосатых мышечных волокон образует сложную систему, в состав к-рой входят четыре основных структурных компонента: 1) сарколемма (sarcolemma), или плазмолемма; 2) система поперечных трубочек (Т-трубочки, каналы Т-с ис те мы, от лат. transversus поперечный); 3) саркоплазматический ретикулум (reticulum sarcoplasmaticum), состоящий из двух отделов — цистер-нового, расположенного в зоне диска I, и канальцевого, составляющего продольно ориентированную сеть (L-канальцы, от лат. longitu-dinalis продольный) в зоне диска А; 4) типичные клеточные мембранные компоненты, входящие в состав пластинчатого комплекса (комплекса Гольджи) и эндоплазматической сети (эргастоплазмы), располагающиеся гл. обр. в околоядерной области (рис. 8). Сарколемма как специфическая оболочка поперечнополосатых мышечных волокон включает два структурных компонента — внутренний, гомогенный, и наружный, волокнистый, по реакциям на гистол, красители близкий к коллагену. При электронно-микроскопическом исследовании выяснилось, что на собственно клеточной, или внутренней, мембране, имеющей типичную для клеточных мембран толщину (ок. 7,5 нм) и структуру, располагается мембрана аморфного строения, содержащая полисахариды, к-рую часто называют базальной мембраной. Базальную мембрану покрывает оболочка, состоящая из тонких коллагеновых волокон. Плазмолемма в зонах телофрагмы образует пальцевидные впячивания (трубочки Т-системы) внутрь волокон между миофибриллами, окружая каждую из них кольцом. По трубочкам Т-системы возбуждение, распространяющееся по плазмолемме, достигает сократительных аппаратов, расположенных внутри волокна. В наиболее тесную связь трубочки Т-системы вступают с цистернами саркоплазматического ретикулума в зоне диска I, где стенки трубочек сближаются со стенками цистерн. На электронограммах профили срезов через области контакта трубочек Т-системы с цистернами саркоплазматического ретикулума имеют вид трех сближенных полостей, называемых триадами.
Цистерны саркоплазматического ретикулума располагаются вокруг миофибрилл в виде муфт, от к-рых вдоль миофибрилл отходят L-канальцы, образующие сеть в зоне мезофрагмы. Вдоль волокон между миофибриллами, занимая большей частью один или два саркомера, располагаются митохондрии (по старой терминологии саркосомы), к к-рым тесно прилежат трубочки Т-системы. Около ядер на электронограммах обнаруживаются типичные цитоплазматические органеллы: пластинчатые комплексы (см. Гольджи комплекс), двойные мембраны эргастоплазмы, рибосомные комплексы, свободные рибосомы (см.), а также иногда комплексы центриолей.
Нервный аппарат поперечнополосатой М. т. состоит из афферентных (чувствительных) нервных структур — нервно-мышечных веретен, а также моторных, или двигательных (эфферентных) структур — моторных пластинок, или бляшек. Нервно-мышечные веретена, инервируются от чувствительных нейронов спинномозговых узлов и представляют собой сложные структуры, в состав к-рых входят специфические измененные (интрафузальные — «внутриверетенные») мышечные волокна, окруженные плотной волокнистой сумкой веретеновидной формы, и иннервирующие их нервные волокна. Моторные бляшки в скелетных мышцах получают нервные импульсы от мотонейронов передних рогов спинного мозга.
Дальнейшее распространение возбуждения по мышечному волокну связано с плазмолеммой и трубочками Т-системы, посредством к-рых возбуждение охватывает внутренние сократительные структуры волокна.
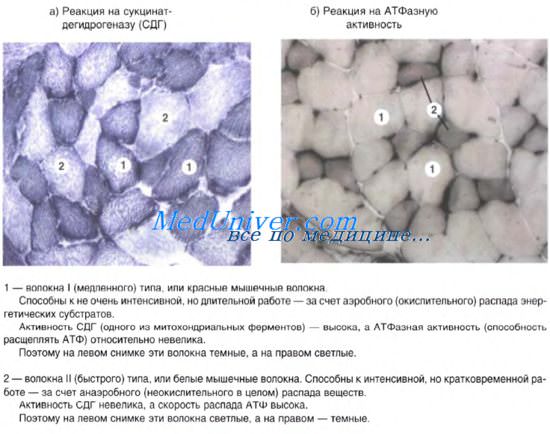
Структурно-функциональные типы мышечных волокон
В скелетной мускулатуре позвоночных, включая млекопитающих и человека, различают два основных типа мышечных волокон: белые, обеспечивающие быструю (фазную) двигательную активность, и красные, способные к длительному неколебательному поддержанию сокращения. Состав почти всех мышц смешанный; в них имеются как белые, так и красные волокна. Преимущественное содержание тех или иных волокон определяет принадлежность мышц к тому или другому типу. Белые мышечные волокна по своей гистол, структуре отличаются высоким содержанием миофибрилл при сравнительно небольшом объеме саркоплазмы, а также более тонкими телофрагмами. Биохимически эти волокна отличаются отсутствием или незначительным содержанием миоглобина, от к-рого зависит окраска мышц. Гистохимически белые мышечные волокна отличаются отсутствием триглицеридов, а также высоким содержанием гликогена и гликолитических ферментов, обеспечивающих энергетические потребности волокна. Для красных мышечных волокон наряду с высоким содержанием миоглобина, обусловливающим их красный цвет, характерно обилие цитоплазматических органелл, в особенности митохондрий в саркоплазме, а также наличие триглицеридов.
Сердечная мышечная ткань (textus muscularis cardiacus)
Гистогенез
Источник развития сердечной М. т. в эмбриогенезе—целомический эпителий, из которого возникает мезенхимный зачаток, подвергающийся дифференцировке в сердечную М. т. Развивающиеся из мезенхимного зачатка миобласты превращаются в сердечные мышечные клетки — кардиомиоциты (myocytus cardiacus), формирующие подобия волокон, в к-рых миобласты не сливаются в мышечные трубочки, а сохраняют клеточную структуру, отделяясь друг от друга вставочными пластинками (см. Сердце).
Строение

Рис. 9. Схема Электронограммы участка сердечной мышцы (продольный срез): 1 — миофибриллы; 2 — диск А; 3 — диск I; 4 — толстые миофиламенты; 5 — тонкие миофиламенты; 6 — митохондрии; 7 — вставочная пластинка; 8 — трубочки Г-системы; 9 — саркоплазматический ретикулум.
В отличие от скелетной сердечная М. т. построена из клеток, связанных в сеть посредством крупных ветвлений, составляющих подобия мышечных волокон. Вследствие такого расположения кардиомиоциты имеют неправильно цилиндрическую форму со ступенчатыми основаниями, к-рые на продольных гистол, срезах имеют вид ступенчатой линии — вставочной пластинки, или вставочного диска (discus intercalatus). В отличие от скелетной М. т. в сердечных мышечных клетках ядра занимают централ ьно-осевое положение, в то время как миофибриллы образуют плотную муфту вокруг ядерной зоны, распространяющуюся до плазмолеммы. При гистол, исследовании с помощью светового микроскопа поперечная исчерченность миофибрилл кажется такой же, как и в миофибриллах скелетной мышечной трубочки. Однако электронно-микроскопическое исследование открывает между ними нек-рые различия, связанные с клеточным строением сердечной М. т. (рис. 9). Т-система в сердечной М. т. развита более интенсивно: трубочки впячиваются вместе с базальной мембраной в саркоплазму. Разделение саркоплазматического ретикулума на цистерно-вый (прилежащий к полудискам I ) и канальцевый (расположенный в зоне диска А) отделы выражено менее значительно, чем в скелетной М. т.: каждый саркомер заключен в муфту из переплетающихся канальцев с небольшими цистернами, к-рые образуют с трубочками Т-системы триады в зоне телофрагм. Характерный признак сердечной М. т.— обилие митохондрий, к-рые локализуются цепочками между миофибриллами, нередко располагаясь в саркомере так, что их концы соприкасаются с телофрагмами. Полости эргастоплазмы, рибосомы и пластинчатые комплексы обнаруживаются гл. обр. около ядер. В связи с клеточным строением сердечной М. т. миофибриллы прерываются в зонах вставочных пластинок, где тонкие (актиновые) нити образуют специфические опорные структуры в виде заостренных арок. Более мощная система Т-тру-бочек и миофибрилл М. т. сердца, возможно, связана с необходимостью обеспечения повышенной прочности сердечной мышцы. По гистол, картине сокращения сердечная М. т. сходна со скелетной. Электронномикроскопическая картина сокращения сердечной М. т. трактуется с позиций теории скольжения, разработанной для скелетной М. т.
Возрастные изменения
Специфическая особенность скелетной М. т. заключается в относительной стабильности ее структурного состава: константное число мышечных волокон в каждой скелетной мышце устанавливается вскоре после рождения и сохраняется до начала старческой инволюции. Возрастные изменения скелетной М. т. характеризуются уменьшением объема мышечных волокон. Те же закономерности, возможно, распространяются и на сердечную М. т., для к-рой также характерны возрастные изменения объема кардиомиоцитов. В связи с отмиранием отдельных мышечных волокон в скелетной М. т. и замещением их соединительной тканью в старческом возрасте уменьшаются эластичность и упругость М. т. Однако в ряде случаев (при правильном гиг. режиме и адекватной мышечной активности) нормальная структура скелетной и сердечной М. т. сохраняется до глубокой старости.
Возрастные изменения гладкой М. т. изучены недостаточно.
Изменения мышечной ткани при патологических состояниях
Гладкая М. т. оболочек органов дыхательной, пищеварительной, мочеполовой систем, а также стенки кровеносных сосудов может подвергаться патол, изменениям гл. обр. в результате нарушения нервной и эндокринной регуляции, а также витаминного, солевого и микроэлементного баланса. Патол. изменение гладкой М. т. (инфильтрация гладких мышечных клеток жировыми и известковыми включениями) наблюдается при атеросклеротическом поражении стенки кровеносных сосудов. Участки гладкой М. т. могут подвергаться малигнизации под влиянием общих этиол, факторов опухолевого роста (см. Опухоли). Встречаются как доброкачественные опухоли — лейомиомы (см.), так и злокачественные — лейомиосаркомы (см.), исходящие из гладкой М. т. ряда органов: кишечника, бронхов и др.
Для скелетной М. т. характерна специфическая структурная реакция на различные нарушения метаболизма: дистрофия миофибриллярного и мембранного аппарата, появление жировых включений, вакуолизация саркоплазмы и особенно гиалиновое перерождение мышечных волокон, выражающееся в развитии в них поперечно расположенных глыбок, полос и узлов (так наз. ценкеровская дегенерация). Денервационная атрофия скелетных мышц на первом этапе характеризуется повышением уровня синтетических процессов (увеличением содержания РНК), усилением пластической активности и структурными изменениями, типичными для регенерационных процессов (переход ядер в центральноосевое положение, развитие миобластов, расщепление мышечных волокон); на более поздних этапах происходит истончение мышечных волокон, ожирение саркоплазмы, дистрофия мышечных волокон с последующим замещением соединительной тканью. Денервационная атрофия скелетных мышц на первом этапе при соответствующих леч. мероприятиях обратима. Постденервацион-ному восстановлению подвергаются гл. обр. моторные бляшки. По данным Р. П. Женевской (1974) нервно-мышечные веретена после длительной денервационной атрофии не восстанавливаются.
Сердечная М. т. реагирует специфическими структурными изменениями на различные патогенные воздействия, включая длительное ги-перфункциональное напряжение, к-рое может вызывать гипертрофию сердечной М. т., выражающуюся гл. обр. в утолщении мышечных волокон. При систематическом отравлении (напр., алкоголем, никотином) наблюдается жировая дистрофия стромы сердечной М. т.
При экспериментальном инфаркте, вызванном перевязкой венечных сосудов, наблюдаются характерные ишемические изменения сердечной М. т.: набухание и разрушение митохондрий, сверхсокращение саркомеров (высокая степень их укорочения) с дезорганизацией протофибриллярного аппарата и саркотубулярной системы. И в скелетной, и в сердечной М. т. могут наблюдаться как доброкачественные (см. Миома, Рабдомиома) так и злокачественные (см. Рабдомиосаркома) новообразования.
Пластическая активность мышечной ткани выражается в процессах первичного (эмбрионального) и вторичного (регенерационного, трансплантационного , ренервационного, гиперфункционального) гистогенеза. Гладкая М. т. обладает достаточно высокими пластическими свойствами, обеспечивающими возможность регенерации при ее повреждениях. Демонстративным примером высокой пластической активности гладкой М. т. является гиперплазия гладкомышечной стенки матки при беременности. В эксперименте доказано, что в этом состоянии гладкая М. т. матки, перенесенная путем аутотрансплантации в измельченном виде на место удаленной скелетной мышцы, может формировать гладкомышечную модель скелетной мышцы. В этом случае мышца состоит из гладких мышечных клеток, образующих по форме подобие скелетной мышцы, связанной посредством сухожилий с определенными точками скелета; сухожилия, очевидно, формируются как фибробластами, так и гладкими мышечными клетками.
Вопрос о способе самообновления гладкой М. т. недостаточно изучен. Школой А. А. Заварзина разработана гипотеза о камбиальном механизме этого процесса (дифференцировка гладкой М. т. происходит за счет особых камбиальных клеток соединительнотканной природы). При изучении регенерации гладкой мышечной ткани в эксперименте наблюдаются митотические деления гладких мышечных клеток.
Как установлено в эксперименте А. Н. Студитским, А. Р. Стригано-вой (1951), А. Н. Студитским (1959, 1978), Р. П. Женевской (1974), регенерационная и трансплантационная активность поперечнополосатой М. т., вопреки старым представлениям о низком уровне ее пластических свойств, оказалась весьма высокой.
Трудность непосредственного применения разработанных в эксперименте методов свободной аутопластики М. т. в хирургической практике заключается в недостаточной изученности видовых пластических свойств скелетной М. т. человека. Тесты, разработанные в эксперименте для испытания пластической активности М. т. (аутотрансплантация измельченной М. т. под кожу, денервация-реиннервация М. т., стимуляция вторичного развития М. т. введением в пищевой рацион тиреоидина, повышающего основной обмен), требуют специального исследования применительно к человеческому организму. Опыт сравнительного (на разных видах животных) изучения свободной аутопластики мышц (либо в измельченном состоянии, либо после предварительной денервации или травматизации) показал, что М. т. собаки обладает высокой пластической активностью. При помощи указанных выше приемов подготовки М. т. у собак удается замещать крупные (до 7 см в длину) фрагменты мышц, что обеспечивает полное восстановление их функции.
По существовавшим ранее представлениям, сердечная М. т. обладает невысокой пластической активностью; доказательством этого положения считалось то, что некротизированные участки сердечной М. т., возникающие в результате инфаркта, замещаются соединительной тканью.
Однако рядом исследователей установлено, что сердечная М. т. обладает специфическим механизмом самовосстановления, к-рый обеспечивается, с одной стороны, соединительнотканным каркасом, замещающим в течение сравнительно короткого срока участок поврежденной сердечной М. т. (напр., омертвевший в результате инфаркта) соединительнотканным рубцом, и более медленно реагирующим на повреждение блоком собственно кардиомиоцитов, за счет к-рых возникает рабочая гипертрофия и гиперплазия сохранившихся участков сердечной мышцы. Существенную роль в самообновлении сердечной М. т. играет внутриклеточная регенерация кардиомиоцитов, включающая постепенную замену функционировавших определенный срок цитоплазматических и ядерных органелл новообразованными (как путем их размножения и разрастания, так и путем молекулярной перестройки).
БИОХИМИЯ МЫШЕЧНОЙ ТКАНИ
Химический состав скелетной мышечной ткани
В скелетной М. т. млекопитающих содержится от 72 до 80% воды, ок. 20—28% веса М. т. составляет сухой остаток, гл. обр. белки. Помимо белков, в состав М. т. входят экстрактивные азотсодержащие вещества, безазотистые вещества (гликоген и другие углеводы, различные липиды, соли органических веществ и др.), а также соли неорганических к-т и другие хим. соединения (табл. 1).
Белки скелетной М. т. делятся на три основные группы: саркоплазматические, миофибриллярные и белки стромы. По данным H. Н. Яковлева (1974), на долю саркоплазматических белков приходится ок. 35%, миофибриллярных — 45% и белков стромы — 20% всего мышечного белка. Указанные группы белков резко отличаются друг от друга по растворимости в воде и солевых средах с различной ионной силой р-ра (полусуммы произведений концентрации каждого иона на квадрат его заряда).
Миофибриллярные белки — миозин, актин и актомиозин растворяются в солевых средах с высокой ионной силой. К миофибриллярным белкам относятся также так наз. регуляторные белки — тропомиозин, тропонин, альфа-актинин, бета-актинин, образующие в мышце с актомиозином единый комплекс.
Главный миофибриллярный белок — миозин составляет 50—55% сухого веса миофибрилл. Работами В. А. Энгельгардта и М. Н. Любимовой показано, что миозин обладает АТФ-азной активностью, т. е. является ферментом со способностью катализировать расщепление АТФ на АДФ и фосфорную к-ту. Хим. энергия АТФ, освобождающаяся в ходе ферментативной реакции, идущей при участии миозина, превращается в механическую энергию сокращающейся мышцы. Относительный мол. вес (масса) миозина скелетных мышц человека ок. 500 000. Молекула миозина, обладающая вытянутой формой (длина ее 150 нм), состоит из двух тяжелых полипептидных цепей с относительным мол. весом 205 000—210 000 и нескольких коротких легких цепей с относительным мол. весом ок. 20 000. Тяжелые цепи образуют длинную закрученную а-спираль («хвост» молекулы), конец каждой тяжелой цепи совместно с легкими цепями создает глобулу («головку» молекулы), способную соединяться с актином. Эти головки выдаются из основного стержня молекулы. Легкие цепи, находящиеся в головке миозиновой молекулы и принимающие участие в проявлении АТФ-азной активности миозина, гетерогенны по аминокислотному составу. Количество легких цепей в молекуле миозина у различных видов животных и в разных типах мышц неодинаково. Молекулы миозина, определенным образом ориентированные в пространстве, образуют так наз. толстые миозиновые нити (толстые миофиламенты) в саркомере.
Актин, составляющий ок. 20% сухого веса миофибрилл, открытый Ф. Штраубом в 1942 г., существует в двух формах: глобулярный актин (Г-актин) и фибриллярный актин (Ф-актин). Молекула Г-актина с относительным мол. весом 42 000 состоит из одной полипептидной цепочки, в образовании к-рой принимают участие 374 аминокислотных остатка. Ф-актин, являющийся продуктом полимеризации Г-актина, имеет структуру двухтяжевой спирали, детали к-рой еще не вполне выяснены.
При мышечном сокращении миозин вступает в соединение с Ф-актином, образуя новый белковый комплекс— актомиозин. Последний обладает АТФ-азной активностью. Однако АТФ-азная активность актомиозина отличается от АТФ-азной активности миозина: актомиозин активируется ионами магния и ингибируется этилендиаминтетраацетатом (ЭДТА) и АТФ в высокой концентрации, тогда как мгозиновая АТФ-аза ингибируется ионами магния, активируется ЭДТА и не ингибируется высокой концентрацией АТФ. Оптимальные значения pH для обеих АТФ-аз также различны.
Содержащиеся в миофибриллах тропомиозин, тропонин и нек-рые другие регуляторные белки непосредственно участвуют в регуляции процесса мышечного сокращения. Молекула тропомиозина, открытого Бейли (К. Bailey) в 1946 г., состоит из двух а-спиралей и имеет вид стержня длиной 40 нм; относительный мол. вес тропомиозина 65 000. На долю тропомиозина приходится ок. 4—7% всех миофпбриллярных белков. Тропонпн — глобулярный белок, открытый Эбаси (S. Ebashi) в 1963 г.; его относительный мол. вес ок. 80 000. В скелетных мышцах человека тропонин составляет лишь ок. 2% всех миофпбриллярных белков.
Тропонин, соединяясь с тропомио-зином, образует комплекс, названный Эбаси нативным тропомиозином. Этот комплекс прикрепляется к актиновым филаментам и придает актомиозину скелетных мышц позвоночных чувствительность к ионам кальция.
Показано, что тропонин способен фосфорилироваться при участии протеинкиназ, зависимых от циклического аденозин-З’,5′-монофосфата (цАМФ). Вопрос о том, имеет ли отношение фосфорилирование тропонина в целостном организме к регуляции мышечного сокращения, остается пока неясным.
Белки стромы в скелетной М. т. представлены в основном коллагеном (см.) и его дериватами, а также эластином (см.). Строма скелетной М. т., остающаяся после исчерпывающей экстракции мышечного гомогената солевыми р-рами с высокой ионной силой, состоит в значительной мере из соединительнотканных элементов стенки сосудов и нервов, а также сарколеммы и нек-рых других структур.
Экстрактивные азотсодержащие вещества скелетной М. т. представлены адениновыми нуклеотидами — АТФ, АДФ и АМФ (см. Аденозинфосфорные кислоты), нуклеотидами неаденинового ряда, креатинфосфатом, креатином, креатинином, карнозином, ансерином, свободными аминокислотами и др. По данным И. И. Иванова (1969), содержание адениновых нуклеотидов в скелетной М. т. кролика (в мкмоль/г сырого веса ткани) составляет: АТФ — 4,43; АДФ — 0,81; АМФ — 0,93. Содержание нуклеотидов неаденинового ряда в М. т. по сравнению с количеством адениновых нуклеотидов очень мало.
На долю азота креатина (см.) и креатинфосфата (см. Фосфагены), по данным Д. Л. Фердмана (1966), приходится до 60% небелкового азота мышц. Креатинфосфат и креатин участвуют в хим. процессах, связанных с мышечным сокращением.
Имидазолсодержащие дипептиды — карнозин (см.) и его метилированное производное ансерин (см.) — способны восстанавливать работоспособность утомленных мышц и влиять на передачу нервных импульсов с нерва на мышцы.
Из свободных аминокислот в М. т. наиболее высока концентрация глутаминовой кислоты (см.) — ок. 120 мг/100 мл и ее амида глутамина (см.) — 80—100 мг/100 мл. В М. т. содержится целый ряд фосфатидов (см.): фосфатидилхолин, фосфатидилэтаноламин, фосфатидил-серин и др.
Эти соединения играют важную роль в структуре М. т., входя в состав клеточных мембран. Фосфатиды принимают участие также в обменных процессах, в частности в качестве субстратов тканевого дыхания. Другие азотсодержащие вещества М. т.: мочевина, мочевая к-та, аденин, гуанин, ксантин и гипоксантин— содержатся в небольшой концентрации и, как правило, являются либо промежуточными, либо конечными продуктами азотистого обмена.
Безазотистые вещества скелетной М. т. представлены в основном гликогеном (см.); его концентрация колеблется от 0,3 до 3% в пересчете на сырой вес. На долю других представителей углеводов приходятся десятые и сотые доли процента. В М. т. находят лишь следы свободной глюкозы и очень мало гексозофосфатов. В процессе метаболизма глюкозы, а также аминокислот в М. т. образуются молочная, Пировиноградная и много других карбоновых к-т. Обнаруживаются также в том или ином количестве нейтральные жиры и холестерин.
Неорганические соли в скелетной М. т. содержатся в виде ионов. Среди катионов наибольшую концентрацию имеют калий (см.) и натрий (см.). Калий гл. обр. сосредоточен внутри мышечных волокон, а натрий находится преимущественно в межклеточном веществе. Значительно ниже в скелетной М. т. содержание магния, кальция и железа; в М. т. содержатся также микроэлементы (см.) — кобальт, алюминий, никель, бор, цинк и др.
Некоторые особенности химического состава гладкой и сердечной мышечной ткани у млекопитающих. Данные о хим. составе гладкой и сердечной М. т. получены в основном на лаб. животных; сведения о хим. составе этих групп М. т. у человека весьма ограничены. Сердечная М. т. по содержанию ряда хим. соединений занимает промежуточное положение между скелетной и гладкой М. т. Так, общее содержание белкового азота в скелетной М. т. кролика— 30—31 мг/г ткани, в миокарде — ок. 23,5 мг/г, а в гладкой мускулатуре матки (миометрий) — в пределах 20,3 мг/г ткани.
Сердечная и особенно гладкая М. т. содержат по сравнению со скелетной М. т. меньше миофпбриллярных белков. Так, содержание миофибрилляр-ных белков (в мг азота на 1 г ткани) в скелетной мускулатуре кролика 17,31, в миокарде — 7,32, а в миометрии — 3,90. Концентрация же белков стромы в миокарде и гладкой М. т. выше, чем в скелетной мускулатуре.
По данным И. И. Иванова (1961), на долю азота белков стромы в скелетной мускулатуре кролика приходится 10,1% от общего азота М. т., в миокарде — 28,2%, а в миометрии — 40,4%. В мышце левого желудочка сердца содержание миофибрилл ярных белков, в частности актомиозина, значительно выше, чем в предсердиях и в ткани миокарда в целом, что, несомненно, связано с более выраженной сократительной функцией этого отдела миокарда. Имеются особенности и во фракционном составе саркоплазматических белков миокарда; так, белков группы миогена в процентном отношении содержится мало, но содержание миоальбумина больше, чем в саркоплазме скелетной мускулатуры.
Содержание АТФ в сердечной М. т. (2,60 мкмоль/г ткани) ниже, чем в скелетной (4,43 мкмоль/г), и выше, чем в гладкой М. т. (1,38 мкмоль/г). Установлено, что содержание АТФ и креатинфосфата неодинаково в различных отделах миокарда. В стенках желудочков сердца, выполняющих значительную работу, этих высокоэргических соединений, обеспечивающих работу мышц энергией, содержится на 40% больше, чем в предсердиях.
По содержанию гликогена, карнозина и ансерина сердечная М. т. также занимает промежуточное положение между скелетной и гладкой М. т. Концентрация имидазол-содержащих дипептидов в миокарде ок. 10 мг/100 мл, в гладкой М. т. обнаруживаются лишь следы ансерина и карнозина.
Имеется определенная зависимость между характером деятельности мышц и содержанием фосфолипидов. Миокард по сравнению со скелетной и гладкой М. т. богаче фосфолипидами (табл. 2), окисление к-рых, невидимому, доставляет значительную часть энергии, необходимой для его сокращения.
Изменение химического состава скелетной мышечной ткани млекопитающих в онтогенезе
Эмбриональная скелетная М. т. по хим. составу значительно отличается от скелетной М. т. взрослых особей. В мышцах эмбрионов содержится больше воды, чем в функционально зрелой М. т. Соответственно общее содержание белка в М. т. эмбрионов (в пересчете на сырую ткань) оказывается более низким, чем в мышцах животных того же вида в постнатальном периоде развития. У куриного эмбриона на 1 г сырой М. т. приходится 10 мг, а у четырех дневных цыплят 32 мг белкового азота. По сравнению с М. т. взрослого организма в функционально незрелой мышце ниже содержание миофибриллярных белков (миозина и актомиозина) и выше — белков стромы, а также миоальбумина и других белков. По мере развития плода содержание миофибриллярных белков повышается и возрастает АТФ-азная активность в мышечных экстрактах.
Для эмбриональной М. т. характерно высокое содержание нуклеопротеидов, а также рибонуклеиновой и дезоксирибонуклеиновой к-т. По мере развития эмбриона содержание нуклеопротеидов и нуклеиновых к-т в М. т. быстро уменьшается. Макроэргических соединений (АТФ и креатинфосфата) в функционально незрелой М. т. значительно меньше, чем в мышцах зрелых особей. Имидазолсодержащие дипептиды (ансерин и карнозин) появляются в составе М. т. в строго определенный период онтогенеза; время их появления тесно связано с началом двигательной функции, формированием рефлекторной дуги, обеспечивающей возможность двигательного рефлекса, появлением кальций-чувствительности актомиозина и началом работы «ионных насосов» (см. Транспорт ионов). Имеются также характерные особенности в ферментных и изоферментных спектрах эмбриональной М. т. Так, установлено, что в ходе онтогенеза изменяется изоферментный спектр лактатдегидрогеназы (см.); в скелетных мышцах 3—5-месячного эмбриона на долю изоферментов лактатдегидрогеназы — ЛДГ3 и ЛДГ2 приходится соответственно 40 и 31% от общей активности лактатдегидрогеназы. В процессе эмбрионального развития в скелетной мускулатуре происходит постепенное возрастание активности изоферментов ЛДГ4 и ЛДГ5 и снижение активности изоферментов ЛДГ1, ЛДГ2 и ЛДГ3, так что у взрослых особей в скелетной мускулатуре наибольшей активностью обладают уже изоферменты ЛДГ5 и ЛДГ4.
Функциональная биохимия поперечнополосатой мышечной ткани
Основной функцией мышц является сокращение. При этом осуществляется работа, связанная с превращением хим. энергии в механическую (см. Механохимические процессы). Мышечное волокно способно сокращаться лишь при наличии в среде АТФ и определенной концентрации ионов кальция. Под влиянием нервного импульса в мышечном волокне происходит изменение проницаемости внутриклеточных мембран и как следствие этого — выход в межфибриллярное пространство из цистерн и трубочек саркоплазматического ретикулума и Т-системы некоторого количества ионов кальция, к-рые связываются с тропонином. В результате создаются условия для взаимодействия актина с миозином с образованием актомиозинового комплекса. При этом за счет энергии АТФ осуществляется скольжение актиновых нитей вдоль миозиновых, т. е. происходит мышечное сокращение (см.). Затем наступает диссоциация актомиозина на миозин и актин, и одновременно начинается новый акт «зарядки» (фосфорилирования) миозина путем взаимодействия его с АТФ в присутствии ионов магния.
Источники энерргии мышечной деятельности
Сократительный аппарат мышечной клетки обеспечивается достаточным количеством энергии в форме АТФ за счет непрерывного ресинтеза этого макроэргического соединения, который происходит прежде всего вследствие трансфосфорилирования АДФ креатинфосфатом. Данная реакция катализируетя ферментом креатинкиназой:

Креатинкиназный путь ресинтеза АТФ является чрезвычайно быстрым и максимально эффективным (за счет каждой молекулы креатинфосфата образуется молекула АТФ). Именно поэтому ряду исследователей не удавалось долгое время установить снижение концентрации АТФ и соответственно повышение концентрации АДФ даже при достаточно продолжительном тетанусе. Лишь применив специфический ингибитор креатинкиназы (1-фтор-2,4-д1 нитрофенол), а также агенты, препятствующие оксидативному превращению АДФ в АТФ, Кейн (D. Cain) и соавт. (1962) смогли продемонстрировать прямой распад АТФ с одновременным приростом неорганического фосфата и АДФ при одиночном сокращении изолированной мышцы лягушки. Эти результаты в дальнейшем были подтверждены рядом других авторов.
Нек-рое количество АТФ может ресинтезироваться в ходе аденилаткиназной (миокиназной) реакции:
![]()
Для любой ткани, в т. ч. и мышечной, имеются два фундаментальных биохим, процесса, в ходе к-рых генерируются богатые энергией фосфорные соединения. Один из этих процессов — гликолиз (см.), другой — тканевое дыхание (см. Окисление биологическое). Наиболее важным и эффективным из них является тканевое дыхание. При достаточном снабжении кислородом мышца, несмотря на анаэробный механизм сокращения, в конечном итоге работает за счет энергии, образующейся в ходе аэробного окисления как продуктов распада углеводов, так и ряда других субстратов тканевого дыхания, в частности жирных к-т, а также ацетата и ацетоацетата (см. Трикарбоновых кислот цикл).
Имеются данные, доказывающие, что креатинфосфат в сердечной М. т. способен выполнять роль не только своего рода депо легко мобилизуемых макроэргршеских фосфатных групп, но и играть также роль транспортной формы макроэргических фосфатных связей, образующихся в процессе тканевого дыхания и связанного с ним окислительного фосфорилирования. По концепции, предложенной В. А. Саксом и др. (1975), перенос энергии из митохондрий в цитоплазму клетки миокарда происходит по следующей схеме: АТФ, образующийся в результате окислительного «фосфорилирования и попадающий в матрикс митохондрий, переносится через внутреннюю мембрану митохондрий с участием специфической АТФ — АДФ-транслоказы на активный центр митохондриального пзофермента креатинкиназы, к-рый расположен на внешней стороне внутренней мембраны; в межмембранном пространстве (в присутствии ионов магния) при наличии в среде креатина образуется равновесный тройной фермент-субстратный комплекс — креатин-креатинкиназа — АТФ — Mg 2+ ; этот комплекс затем распадается с образованием креатинфосфата и АДФ — Mg 2+ . Креатинфосфат диффундирует в цитоплазму, где используется в миофибриллярной креатинкиназной реакции для рефосфорилирования АДФ, образовавшегося в акте сокращения. Высказываются предположения, что не только в сердечной, но и в скелетной М. т. имеет место подобный путь транспорта энергии из митохондрий в миофибриллы.
При умеренной работе мышца может покрывать свои энергетические затраты за счет аэробного метаболизма. Однако при больших нагрузках, когда возможность снабжения кислородом отстает от потребности в нем, в мышцах используется гл политический путь снабжения энергией. При интенсивной мышечной работе скорость расщепления гликогена или глюкозы с образованием молочной к-ты увеличивается в сотни раз. Соответственно содержание молочной к-ты в М. т. может повышаться до 100—120 мг в 100 мл и выше. Молочная к-та с током крови в значительном количестве поступает в печень, где за счет энергии окислительных процессов ресинтезируется в глюкозу и гликоген (глюконеогенез).
Перечисленные механизмы ресинтеза АТФ при мышечной деятельности включаются в строго определенной последовательности. Наиболее экстренным является креатинкиназный механизм, и лишь после — 20 сек. интенсивной работы начинается усиление гликолиза, этот процесс достигает максимума через 40—80 сек. При менее интенсивной работе большее значение приобретает аэробный путь ресинтеза АТФ.
Содержание АТФ и креатинфосфата в сердечной М. т. ниже, чем в скелетной мускулатуре, а расход АТФ весьма велик, поэтому ресинтез АТФ в миокарде проходит намного интенсивнее, чем в скелетной М. т. Для сердечной мышцы теплокровных животных и человека основным путем образования богатых энергией фосфорных соединений является путь окислительного фосфорилирования (см.), связанный с поглощением кислорода. Регенерация АТФ в процессе анаэробного расщепления углеводов (гликолиз) в сердце человека практического значения не имеет. Именно поэтому сердечная мышца очень чувствительна к недостатку кислорода. Характерной особенностью обмена веществ сердечной М. т. по сравнению со скелетной является также и то, что аэробное окисление веществ неуглеводной природы при работе сердечной мышцы имеет большее значение, чем при сокращении скелетной мышцы. Только 30—35% кислорода, поглощаемого сердцем в норме, расходуется на окисление углеводов и продуктов их превращения. Главным же субстратом дыхания в сердечной мышце являются жирные к-ты. Окисление неуглеводных веществ в норме обеспечивает ок. 65—70% потребности миокарда в энергии. Однако при повышении уровня углеводов в крови (напр., сразу после еды) потребление их сердечной мышцей увеличивается. Расход кислорода на окисление углеводов резко возрастает, а скорость окисления жирных к-т в миокарде снижается. Наоборот, натощак и при голодании энергетические потребности сердца в основном покрываются за счет окисления жирных к-т.
Биохимические изменения в скелетной мышечной ткани при патологии
Общим для большинства заболеваний мышц (прогрессирующие мышечные дистрофии, атрофия мышц на почве их денервации, полимиозит, поражения мышц при нек-рых авитаминозах и др.) является резкое снижение в мышцах содержания миофибриллярных белков, возрастание концентрации белков стромы и нек-рых саркоплазматических белков, в т. ч. миоальбу-мина. Наряду с изменением фракционного состава мышечных белков при поражениях мышц наблюдается снижение концентрации АТФ и креатинфосфата. Отмечается также снижение АТФ-азной активности сократительных белков (миозина), уменьшение содержания имидазол-содержащих дипептидов. Считают, что снижение содержания ансерина и карнозина связано! не с нарушением биосинтетических процессов, а с усилением распада дипептидов.
При прогрессирующих мышечных дистрофиях и других заболеваниях мышц, связанных с распадом мышечной ткани, часто отмечаются сдвиги в фосфолипидном составе мышц: значительно снижается содержание фосфатидилхолина и фосфатидилэтаноламина, концентрация же сфингомиелина и лизофосфатидилхолина повышается. Механизмы изменения фосфолипидного состава М. т. при патологии еще не выяснены, неизвестна также роль этих сдвигов в патогенезе мышечных дистрофий.
Для многих форм патологии М. т. характерно нарушение метаболизма креатина и его усиленное выделение с мочой — креатинурия (см.). Вопрос о причинах креатинурии при заболеваниях мышц не может еще считаться окончательно решенным. Креатинурия у больных миопатией, возможно, является результатом нарушения в скелетной М. т. процессов фиксации (удержания) креатина и его фосфорилирования. В результате нарушения процесса синтеза креатинфосфата не образуется и креатинин; содержание последнего в моче резко снижается. Вследствие креатинурии и нарушения синтеза креатинина резко повышается креатининовый коэффициент (см. Креатин).
При патологии М. т. можно наблюдать определенную закономерность в изменении активности ферментов в мышцах: уменьшается активность энзимов, локализованных в саркоплазме; незначительно изменяется активность ферментов, связанных с митохондриями; заметно возрастает активность лизосомальных ферментов. Наконец, показано, что при многих заболеваниях мышечной системы наступают сдвиги в системе циклического аденозинмонофосфата (цАМФ): в М. т. снижается содержание цАМФ, повышается активность фосфодиэстеразы, и нарушается способность аденилатциклазы активироваться под влиянием адреналина и фторида натрия.
Таблицы
Таблнца 1. Химический состав скелетной мышечной ткани млекопитающих (по И. И. Иванову, 1974)
Гладкая мышечная ткань мочевого пузыря

1 — гладкие миоциты (клетки, образующие гладкую мышечную ткань): имеют веретиновидную форму и часто объединяются в пучки. Толстые и тонкие миофиламенты не формируют миофибриллы, поэтому клетки лишены поперечной исчерченности. Прочие внутриклеточные структуры: 2 — ядро: по форме — палочковидное, расположено в центре клетки; 3 — гранулярная ЭПС (эндоплазматическая сеть): участвует в синтезе компонентов межклеточного вещества — протеогликанов и др.
Контакты и окружение клеток: 4 — нексусы: контакты, соединяющие соседние миоциты в пучке; 5 — базальная мембрана: окружает каждый гладкий миоцит; 6— эндомизий: прослойки соединительной ткани вокруг миоцитов.
Развивается из мезенхимы, иннервируется вегетативной НС, медленно, но долго сокращается.
Мышечная ткань соматического типа (языка кролика) – поперечно-полосатая скелетная
Малое увеличение Большое увеличение



а) Малое увеличение 1 — продольно срезанные пучки мышечных волокон; 2 — поперечно срезанные пучки мышечных волокон; 3 — эндомизий: прослойки рыхлой соединительной ткани между мышечными волокнами; 4 — перимизий: прослойка рыхлой соединительной ткани между пучками мышечных волокон.

б) Среднее увеличение
У продольно срезанных волокон наблюдаются признаки, характерные для скелетной мышечной ткани. 4 — ядра: в волокне (симпласте) их много, а располагаются они на периферии волокна, непосредственно под сарколеммой; 5 и 6 — темные и светлые полоски в волокнах; их чередование придает волокнам поперечную исчерченность.

в) Большое увеличение
7 — миофибриллы: на поперечном сечении мышечного волокна имеют вид точек и заполняют почти все сечение волокна.

Окраска железным гематоксилином 1 — ядра: занимают периферическое положение в волокнах. Видна поперечная исчерченность волокон.
Сердечная поперечно-полосатая мышечная ткань


Большое увеличение

1 — поперечная исчерченность в функциональных волокнах миокарда. 2 — вставочные диски: имеют вид темных полос и разделяют функциональные волокна на отдельные клетки — кардиомиоциты – клетки цилиндрической формы. В кардиомиоцитах: миофибриллы(сакромеры+З-линия), ядра, Т-трубочки(вокруг миобирилл), Л-система. В клетках ядра занимают центральное положение.
В области вставочных дисков, м/у кардиомиоцитами существует несколько видов контактов: интердигитации-пальцевидные впячивания клеток друг в друга, десмосомы- обеспечивают более прочное соединение, нексусы- электрическая связь.
7. Безмиелиновые нервные волокна


(по Т.Н.Радостиной, Ю.И.Афанасьеву, Т. С. Румянцевой)
1 — ядро леммоцита (шванновской клетки): располагается в центре волокна;
2 — осевые цилиндры (отростки нейронов): 10—20 осевых цилиндров погружено по периферии волокна в цитоплазму леммоцита. Над каждым цилиндром плазмолемма леммоцита смыкается — так, что образуется «брыжейка», или
4 — базальная мембрана вокруг нервного волокна.


Подпись к обоим снимкам. 1 — нервные волокна. Они отделены друг от друга в процессе приготовления препарата; 2 — ядра олигодендроцитов (леммоцитов, или шванновских клеток): узкие, расположены в центре волокна и ориентированы по его оси.
Поперечнополосатая мышечная ткань
Различают поперечнополосатую скелетную и сердечную мышечные ткани.
Поперечнополосатая скелетная мышечная ткань
Состоит из продольно ориентированных мышечных волокон (симпласты). Длина волокон от 1 до 40 мм, диаметр – 0,1 мм.
Строение мышечного волокна
Мышечное волокно имеет оболочку – сарколемму, состоящую из:
сети ретикулярных волокон
Ядра овальной формы, количеством несколько сот на 1 см длины, располагаются под сарколеммой. Средняя часть волокна заполнена миофибриллами (сократительный аппарат). Жидкая часть цитоплазмы волокна – саркоплазма, заполняет промежутки между миофибриллами и содержит белок – миоглобин (Рис 12). Между миофибриллами также располагается большое количество продольно ориентированных митохондрий (саркосом). Много гранул гликогена.

В зависимости от соотношения миофибрилл и саркоплазмы, различают следующие виды мышц: белые и красные. В белых мышцах мало саркоплазмы, но много миофибрилл. Сокращения их быстрые, кратковременные; они быстро «утомляются». В красных мышцах много саркоплазмы и митохондрий, но меньше миофибрилл. Сокращения их медленные, но длительные; они медленно «утомляются». Примеры:
У домашних птиц грудные мышцы – белые, а мышцы ног – красные, по причине чего они не способны к длительному полету;
У диких птиц грудные мышцы – красные, благодаря чему они способны к длительному полету.
У человека промежуточный тип мышц, т.е. примерно одинаковые соотношения миофибрилл и саркоплазмы.
Ультрамикроскопическое строение миофибрилл
Миофибриллы состоят из двух чередующих сегментов, которые называются:
Диск А (темный диск) – от слова «анизотропный» и
Диск I(светлый диск) – от слова «изотропный»
В средней части диска I проходит полоска Z, а средняя часть диска А просветленная и носит название – зона Н. Участок миофибрилл между двумя соседними Z – полосками – саркомер.
Формула саркомера, т.е. саркомер состоит из: ½ I + A + ½ I. Саркомер – это элементарная сократительная единица миофибрилл (Рис. 13).
Под электронным микроскопом можно видеть, что миофибриллы состоят из протофибрилл. Различают два вида протофибрилл:
толстые – диаметр 100 Å (белок миозини)
тонкие – диаметр 50 Å (белок актин).
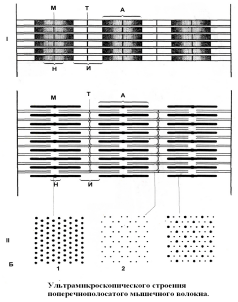
Толстые прото-фибриллы занимают диски А, а тонкие диски I. Концы толстых протофибрилл ни к чему не прикреплены, лежат свободно. Тонкие протофибриллы одним концом прикреплены к полоске – Z, а другой конец лежит свободно в промежутках между толстыми протофибриллами на периферии дисков А. Таким образом, периферические (более темные) части дисков А состоят из толстых и тонких протофибрилл, а средние (более светлые) части состоят только из толстых протофибрилл.
Периферические части дисков А, состоящие из толстых и тонких протофибрилл, носят название – зоны перекрытия. В зонах перекрытия толстые и тонкие протофибриллы располагаются строго определенным образом: каждая толстая протофибрилла окружена шестью тонкими и при их соединении линиями образуется фигура шестиугольника, отсюда и название их взаимного расположения: гексагональная упаковка протофибрилл.